
矮柱兰Thelasis pygmaea (Griff.) Bl.
矮柱兰Thelasis pygmaea (Griff.) Bl.
1. 矮柱兰(中国高等植物图鉴)闭花八粉兰(中山自然科学大辞典)图版8: 1-3
Thelasis pygmaea (Griff.) Bl., Fl. Jav. Orch. : 23. 1858; Lindl. in J. Linn. Soc. Bot. 3: 63. 1859; Hance in J. Linn. Soc. Bot. 13: 127. 1872; Hook. f., Fl. Brit. Ind. 6: 86. 1890; King et Pantl. in Ann. Bot. Gard. Calcutta 8: 249. pl. 331. 1898; Seidenf. in Opera Bot. 89: 156. fig. 101. 1986, et 114: 193. 1992; 台湾兰科植物彩色图鉴 2: 323, 663, 1990; Z. H. Tsi et S. C. Chen in Acta Phytotax. Sin. 33 (3): 296. 1995; Z. H. Tsi et al., l. c. 33 (6): 591. 1995. ——Euproboscis Pygmaea Griff. in Calc. J. Nat. Hist. 5: 731. t. 26. 1845. ——Thelasis triptera Rchb. f. in Bonplandia 3: 219. 1853; 中国高等植物图鉴 5: 750. 图8329. 1976; 海南植物志 4: 248. 图1120. 1977; 台湾兰科植物3: 272-273 (图). 1987. ——T. elongata Bl., Bot. Mus. Lugd. 2: 187. 1856. ——T. hongkongensis Rolfe in Kew Bull. : 199. 1896, et in J. Linn. Soc. Bot. 36: 39. 1903; S. Y. Hu, Gen. Orch. Hong Kong: fig. 39. 1977. ——T. clausa Fukuyama in Bot. Mag. Tokyo 49: 440. 1935; Masamune in J. Geobot. 21 (1): t. 187. 1973; 台湾兰科植物彩色图鉴 1: 504. 1977; 台湾植物志 5: 1109. 1978.
假鳞茎聚生,扁球形(上下压扁),高3-10毫米,粗7-18毫米,顶端通常具1枚大叶和1 (-2) 枚小叶。大叶狭长圆状倒披针形至近狭长圆形,长 (2.7-) 4-8 (-12) 厘米,宽6-13毫米,先端钝、急尖或不等的2裂,稍肉质,基部收狭成短柄并内卷;小叶近长圆形,长0.7-1.5厘米,较少不存在。花葶生于假鳞茎基部,高10-20 (-30) 厘米,纤细,有2-3枚抱茎的鞘;总状花序初期较短,长1-2厘米,随着花的开放,逐渐延长,可达5-10厘米并多少外弯或下弯,生有许多密集的小花;花序轴常较肥厚;花苞片卵状三角形或卵状披针形,长约2毫米,宿存,常稍呈紫色;花梗和子房长约3毫米;花黄绿色,平展,不甚张开;中萼片卵状披针形或长圆状披针形,长2-2.5毫米,下部宽约1毫米;侧萼片与中萼片相似,但背面具龙骨状突起或有时呈狭翅状;花瓣近长圆形或狭长圆形,长约2毫米,宽约0.7毫米;唇瓣卵状三角形,长约1.8毫米,基部宽约1.2毫米,先端渐尖,边缘内卷;蕊柱短,无蕊柱足;花药狭卵形,直立,长约0.7毫米;蕊喙较长,直立,长达1.2毫米。花期4-10月。
产台湾、海南和云南南部至东南部(景洪、勐腊、富宁)。生于溪谷旁树干上、山崖树枝上或林中石上,海拔1100米以下。尼泊尔、锡金、印度、缅甸、泰国、越南、马来西亚、印度尼西亚和菲律宾也有分布。模式标本采自菲律宾。
形态特征:假鳞茎聚生,扁球形(上下压扁),高3—10毫米,粗7—18毫米,顶端通常具1枚大叶和1(--2)枚小叶。大叶狭长圆状倒披针形至近狭长圆形,长(2.7--)4—8(一12)厘米,宽6—13毫米,先端钝、急尖或不等的2裂,稍肉质,基部收狭成短柄并内卷;小叶近长圆形,长0.7—1.5厘米,较少不存在。花葶生于假鳞茎基部,高10—20(--30)厘米,纤细,有2—3枚抱茎的鞘;总状花序初期较短,长1—2厘米,随着花的开放,逐渐延长,可达5--10厘米并多少外弯或下弯,生有许多密集的小花;花序轴常较肥厚;花苞片卵状三角形或卵状披针形,长约2毫米,宿存,常稍呈紫色;花梗和子房长约3毫米;花黄绿色,平展,不甚张开;中萼片卵状披针形或长圆状披针形,长2—2.5毫米,下部宽约1毫米;侧萼片与中萼片相似,但背面具龙骨状突起或有时呈狭翅状;花瓣近长圆形或狭长圆形,长约2毫米,宽约0.7毫米;唇瓣卵状三角形,长约1.8毫米,基部宽约1.2毫米,先端渐尖,边缘内卷;蕊柱短,无蕊柱足;花药狭卵形,直立,长约0.7毫米;蕊喙较长,直立,长达1.2毫米。花期4—10月。
产地分布:产台湾、海南和云南南部至东南部(景洪、勐腊、富宁)。生于溪谷旁树干上、山崖树枝上或林中石上,海拔1100米以下。尼泊尔、锡金、印度、缅甸、泰国、越南、马来西亚、印度尼西亚和菲律宾也有分布。模式标本采自菲律宾。
参考文献:Thelasis pygmaea(Griff.)Bl,F1.Jav.Orch.:23.1858;Lindl.In J.Linn.Soc.Bot.3:63.1859;Hance in J.Linn.Soc.Bot.13:127.1872;Hook.F., F1.Brit.Ind.6:86.1890;King et Pantl.In Ann.Bot.Gard.Calcutta8:249.P1. 331.1898;Seidenf.In Opera Bot.89:156.Fig.101.1986,et 114:193.1992;台 湾兰科植物彩色图鉴2:323,663,1990;2.H.TsietS.C.Chen in Acta Phytotax. Sin.33(3):296.1995;Z.H.Tsi et al,1,c.33(6):591.1995.——Eupro boscis Pygmaea Griff.In Calc.J.Nat.Hist.5:731.T.26,1845.——Thelasis triptera Rchb.F.In Bonplandia 3:219.1853;中国高等植物图鉴5:750. 图8329, 1976;海南植物志4:248.图1120.1977;台湾兰科植物3:272--273(图). 1987.——T.Elongata Bl,Bot.Mus.Lugd.2:187.1856.——T.honZAongen— sis Rolfe in Kew Bull.:199.1896,et in J.Linn.Soc.Bot.36:39.1903;S.Y, Hu,Gen.Orch.Hong Kong:fig.39.1977.——T.Clatisa Fukuyama inBot.Mag, Toky049:440.1935;Masamune in J.Geobot.21(1):t.187. 1973;台湾兰科植物 彩色图鉴1:504.1977;台湾植物志5:1109.1978.
本属概述:Bl,Bijdr.8:385.1825;Seidenf.In Opera Bot.89:154. 1986.——Euproboscis Griff. In Calc. J. Nat. Hist. 5:371. 1845.附生小草本,具假鳞茎或缩短的茎,后者常包藏于套叠的叶鞘中。叶1—2枚,生 于假鳞茎顶端或多枚二列地着生于缩短的茎上,后者基部具鞘并互相套迭,有时有关 节。花葶侧生于假鳞茎或短茎基部,通常较细长;总状花序或穗状花序具多花;花很 小,几乎不张开;萼片相似,靠合,仅先端分离;侧萼片背面常有龙骨状突起;花瓣略 小于萼片;唇瓣不裂,多少凹陷,着生于蕊柱基部;蕊柱短,无蕊柱足;花药直立,药 帽先端收狭为钻状;花粉团8个,每4个为一群,蜡质,共同连接于一个细长而上部稍 扩大的花粉团柄上;粘盘近狭椭圆形;蕊喙顶生,直立,渐尖,2裂(粘盘脱出后);柱 头较大。蒴果较小。本属模式种:Thelasis obtusa Bl.全属约20种,产亚洲热带地区,主要见于东南亚,向北可到尼泊尔和我国南部, 向南可到新几内亚岛。我国有2种。
亚族概述:Schltr,Orch.340.1914,etinNotizbl.Bot.Gard.Berlin9:576.1926."Thelasieae",附生草本,具或不具假鳞茎,有时有茎;茎一般纤细,长或短。叶茎生或生于假鳞茎顶端,扁平,对折,革质或近肉质,有时两侧压扁或近圆柱形,具或不具关节。花葶侧生;总状花序具少数或多数花;花一般较小,常不甚张开;萼片与花瓣离生,有时靠合;唇瓣生于蕊柱基部或短的蕊柱足上;蕊柱一般较短,具或不具蕊柱足;花药生于蕊柱顶端背侧;花粉团8或4个,蜡质,具有一个共同的花粉团柄,有粘盘;柱头1个,凹陷。本亚族模式属:Thelasis Bl.共含6属,分布于亚洲热带地区和大洋洲。我国有2属。
本族概述:Humb,Bonpl.EtKunth,Nov.Gen.Sp.Pl.1:269.1815.附生或较少地生或腐生草本,偶见攀援藤本;附生种类常有假鳞茎或肉质茎。叶基生或茎生,常为革质、纸质或肉质,罕有草质,少数无叶(腐生种类)。花变化较大;萼片合生或离生;唇瓣有距或无距;花药以狭窄基部或花丝连接于蕊柱,常俯倾,两药室一般密接并藏于药帽之内,花后常枯萎或脱落;花粉团一般4—8个,较少2个,蜡质或有时粒粉质,具或不具花粉团柄或粘盘,偶见粘盘柄;蕊喙一般位于柱头上方。本族模式属:EpidendrumL.共约340属,全球均有分布,主要见于热带。我国有29个亚族含72属。
亚科概述:地生、附生或腐生草本,具根状茎、块茎或假鳞茎。叶茎生或基生,折扇状、对折或具网状脉,草质至革质。花序(或花葶)顶生或侧生;花通常两侧对称,有特化的唇瓣,少有例外,一般扭转,具1室子房;蕊柱通常具1枚能育雄蕊;花粉粘合成团块,称花粉团;花粉团2--8个,粒粉质或蜡质,其一端常伸出花粉团柄并连接于粘盘,也有直接连接于粘盘或通过粘盘柄连接于粘盘的,较少既无花粉团柄又无粘盘柄与粘盘的;柱头1或2,凹陷或凸出;蕊喙通常存在,大或小。果实通常为蒴果;较少呈荚果状。种子通常两端有延长的翅,较少无翅并有厚的外种皮。本亚科模式属:OrchisL.共约690余属,全球均有分布。我国有4个族,42个亚族共167属。
本科概述:地生、附生或较少为腐生草本,极罕为攀援藤本;地生与腐生种类常有块茎或肥厚的根状茎,附生种类常有由茎的一部分膨大而成的肉质假鳞茎。叶基生或茎生,后者通常互生或生于假鳞茎顶端或近顶端处,扁平或有时圆柱形或两侧压扁,基部具或不具关节。花葶或花序顶生或侧生;花常排列成总状花序或圆锥花序,少有为缩短的头状花序或减退为单花,两性,通常两侧对称;花被片6,2轮;萼片离生或不同程度的合生;中央1枚花瓣的形态常有较大的特化,明显不同于2枚侧生花瓣,称唇瓣,唇瓣由于花(花梗和子房)作180°扭转或90°弯曲,常处于下方(远轴的一方);子房下位,1室,侧膜胎座,较少3室而具中轴胎座;除子房外整个雌雄蕊器官完全融合成柱状体,称蕊柱;蕊柱顶端一般具药床和1个花药,腹面有1个柱头穴,柱头与花药之间有1个舌状器官,称蕊喙(源自柱头上裂片),极罕具2—3枚花药(雄蕊)、2个隆起的柱头或不具蕊喙的;蕊柱基部有时向前下方延伸成足状,称蕊柱足,此时2枚侧萼片基部常着生于蕊柱足上,形成囊状结构,称萼囊;花粉通常粘合成团块,称花粉团,花粉团的一端常变成柄状物,称花粉团柄;花粉团柄连接于由蕊喙的一部分变成固态粘块即粘盘上,有时粘盘还有柄状附属物,称粘盘柄;花粉团、花粉团柄、粘盘柄和粘盘连接在一起,称花粉块,但有的花粉块不具花粉团柄或粘盘柄,有的不具粘盘而只有粘质团。果实通常为蒴果,较少呈荚果状,具极多种子。种子细小,无胚乳,种皮常在两端延长成翅状。科的模式属:OrchisL.全科约有700属20000种,产全球热带地区和亚热带地区,少数种类也见于温带地区。我国有171属1247种以及许多亚种、变种和变型。兰科是一个进化而复杂的科,至今对科内各类群间亲缘关系的了解仍然是十分初步的。在目前已发表的许多兰科系统中,以R.Dressier(1981,1993)系统较为全面,但该系统对鸟巢兰族(ridb.Neottieae)的处理是有欠缺的。幸而C.Seidenfaden等人在“TheOrehidsofIndochina”(OperaBot.114:5--502.1992)和“TheOrehidsofPeninsularMalaysiaandSingapore”(1992)中有较合理的处理。本书基本上采用了后者的框架,但将等级作了调整,而对亚族的组成则更多地参考Dressler系统(PhylogenyandClassifica-tionoftheOn:hidFamily,1993)。本书将兰科分为3个亚科,兰亚科之下分4个族。其中兰族(Orchideae)与万代兰族(Vandeae)是比较自然的类群,而鸟巢兰族(Neottieae)与树兰族(Epidendreae)则尚存在不少问题。在许多近代兰科系统中,上述的4个族均被作为亚科,在亚科之下又有大量的族,在族之下则是亚族。应当说,大多数亚族的建立是比较客观的。但族的组合则含有相当多的主观性,甚至是勉强的拼凑。因此,本书避繁就简,在兰亚科之下只承认4个族,族之下则包含较多的亚族。这样不仅简化了层次,而且更为客观和实用。当然,这也只是权宜之计,有些问题仍然未能解决,如鸟巢兰族与树兰族的特征交叉过多,有些亚族较为庞杂等。这里就不作详细讨论了。这里要说明的是,本书所提供的“亚科与族的检索表”和“兰亚科分亚族检索表”均系根据国产种类编写而成的,未必完全符合族与亚族的全面特征,在使用时务请加以注意。
亚科概述:地生、附生或腐生草本,具根状茎、块茎或假鳞茎。叶茎生或基生,折扇状、对折或具网状脉,草质至革质。花序(或花葶)顶生或侧生;花通常两侧对称,有特化的唇瓣,少有例外,一般扭转,具1室子房;蕊柱通常具1枚能育雄蕊;花粉粘合成团块,称花粉团;花粉团2--8个,粒粉质或蜡质,其一端常伸出花粉团柄并连接于粘盘,也有直接连接于粘盘或通过粘盘柄连接于粘盘的,较少既无花粉团柄又无粘盘柄与粘盘的;柱头1或2,凹陷或凸出;蕊喙通常存在,大或小。果实通常为蒴果;较少呈荚果状。种子通常两端有延长的翅,较少无翅并有厚的外种皮。本亚科模式属:OrchisL.共约690余属,全球均有分布。我国有4个族,42个亚族共167属。
本科概述:地生、附生或较少为腐生草本,极罕为攀援藤本;地生与腐生种类常有块茎或肥厚的根状茎,附生种类常有由茎的一部分膨大而成的肉质假鳞茎。叶基生或茎生,后者通常互生或生于假鳞茎顶端或近顶端处,扁平或有时圆柱形或两侧压扁,基部具或不具关节。花葶或花序顶生或侧生;花常排列成总状花序或圆锥花序,少有为缩短的头状花序或减退为单花,两性,通常两侧对称;花被片6,2轮;萼片离生或不同程度的合生;中央1枚花瓣的形态常有较大的特化,明显不同于2枚侧生花瓣,称唇瓣,唇瓣由于花(花梗和子房)作180°扭转或90°弯曲,常处于下方(远轴的一方);子房下位,1室,侧膜胎座,较少3室而具中轴胎座;除子房外整个雌雄蕊器官完全融合成柱状体,称蕊柱;蕊柱顶端一般具药床和1个花药,腹面有1个柱头穴,柱头与花药之间有1个舌状器官,称蕊喙(源自柱头上裂片),极罕具2—3枚花药(雄蕊)、2个隆起的柱头或不具蕊喙的;蕊柱基部有时向前下方延伸成足状,称蕊柱足,此时2枚侧萼片基部常着生于蕊柱足上,形成囊状结构,称萼囊;花粉通常粘合成团块,称花粉团,花粉团的一端常变成柄状物,称花粉团柄;花粉团柄连接于由蕊喙的一部分变成固态粘块即粘盘上,有时粘盘还有柄状附属物,称粘盘柄;花粉团、花粉团柄、粘盘柄和粘盘连接在一起,称花粉块,但有的花粉块不具花粉团柄或粘盘柄,有的不具粘盘而只有粘质团。果实通常为蒴果,较少呈荚果状,具极多种子。种子细小,无胚乳,种皮常在两端延长成翅状。科的模式属:OrchisL.全科约有700属20000种,产全球热带地区和亚热带地区,少数种类也见于温带地区。我国有171属1247种以及许多亚种、变种和变型。兰科是一个进化而复杂的科,至今对科内各类群间亲缘关系的了解仍然是十分初步的。在目前已发表的许多兰科系统中,以R.Dressier(1981,1993)系统较为全面,但该系统对鸟巢兰族(ridb.Neottieae)的处理是有欠缺的。幸而C.Seidenfaden等人在“TheOrehidsofIndochina”(OperaBot.114:5--502.1992)和“TheOrehidsofPeninsularMalaysiaandSingapore”(1992)中有较合理的处理。本书基本上采用了后者的框架,但将等级作了调整,而对亚族的组成则更多地参考Dressler系统(PhylogenyandClassifica-tionoftheOn:hidFamily,1993)。本书将兰科分为3个亚科,兰亚科之下分4个族。其中兰族(Orchideae)与万代兰族(Vandeae)是比较自然的类群,而鸟巢兰族(Neottieae)与树兰族(Epidendreae)则尚存在不少问题。在许多近代兰科系统中,上述的4个族均被作为亚科,在亚科之下又有大量的族,在族之下则是亚族。应当说,大多数亚族的建立是比较客观的。但族的组合则含有相当多的主观性,甚至是勉强的拼凑。因此,本书避繁就简,在兰亚科之下只承认4个族,族之下则包含较多的亚族。这样不仅简化了层次,而且更为客观和实用。当然,这也只是权宜之计,有些问题仍然未能解决,如鸟巢兰族与树兰族的特征交叉过多,有些亚族较为庞杂等。这里就不作详细讨论了。这里要说明的是,本书所提供的“亚科与族的检索表”和“兰亚科分亚族检索表”均系根据国产种类编写而成的,未必完全符合族与亚族的全面特征,在使用时务请加以注意。
《Flora of China》 Vol. 25 (2009)
Thelasis pygmaea
矮柱兰
别名:闭花八粉兰;
科名:兰科 Orchidaceae
属名:矮柱兰属 Thelasis
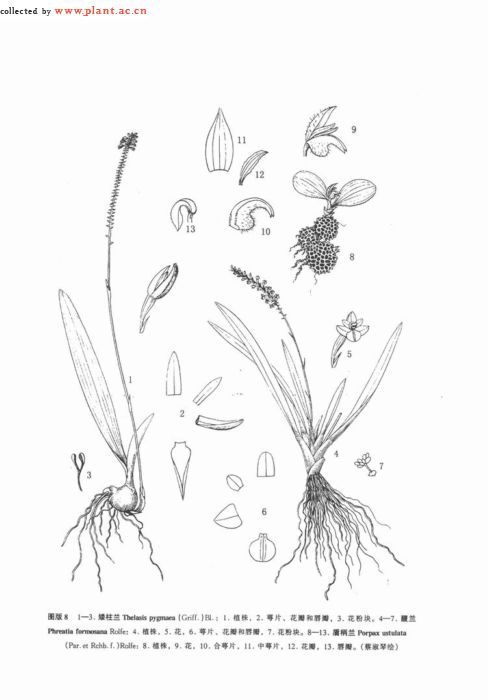
1. 矮柱兰(中国高等植物图鉴)闭花八粉兰(中山自然科学大辞典)图版8: 1-3
Thelasis pygmaea (Griff.) Bl., Fl. Jav. Orch. : 23. 1858; Lindl. in J. Linn. Soc. Bot. 3: 63. 1859; Hance in J. Linn. Soc. Bot. 13: 127. 1872; Hook. f., Fl. Brit. Ind. 6: 86. 1890; King et Pantl. in Ann. Bot. Gard. Calcutta 8: 249. pl. 331. 1898; Seidenf. in Opera Bot. 89: 156. fig. 101. 1986, et 114: 193. 1992; 台湾兰科植物彩色图鉴 2: 323, 663, 1990; Z. H. Tsi et S. C. Chen in Acta Phytotax. Sin. 33 (3): 296. 1995; Z. H. Tsi et al., l. c. 33 (6): 591. 1995. ——Euproboscis Pygmaea Griff. in Calc. J. Nat. Hist. 5: 731. t. 26. 1845. ——Thelasis triptera Rchb. f. in Bonplandia 3: 219. 1853; 中国高等植物图鉴 5: 750. 图8329. 1976; 海南植物志 4: 248. 图1120. 1977; 台湾兰科植物3: 272-273 (图). 1987. ——T. elongata Bl., Bot. Mus. Lugd. 2: 187. 1856. ——T. hongkongensis Rolfe in Kew Bull. : 199. 1896, et in J. Linn. Soc. Bot. 36: 39. 1903; S. Y. Hu, Gen. Orch. Hong Kong: fig. 39. 1977. ——T. clausa Fukuyama in Bot. Mag. Tokyo 49: 440. 1935; Masamune in J. Geobot. 21 (1): t. 187. 1973; 台湾兰科植物彩色图鉴 1: 504. 1977; 台湾植物志 5: 1109. 1978.
假鳞茎聚生,扁球形(上下压扁),高3-10毫米,粗7-18毫米,顶端通常具1枚大叶和1 (-2) 枚小叶。大叶狭长圆状倒披针形至近狭长圆形,长 (2.7-) 4-8 (-12) 厘米,宽6-13毫米,先端钝、急尖或不等的2裂,稍肉质,基部收狭成短柄并内卷;小叶近长圆形,长0.7-1.5厘米,较少不存在。花葶生于假鳞茎基部,高10-20 (-30) 厘米,纤细,有2-3枚抱茎的鞘;总状花序初期较短,长1-2厘米,随着花的开放,逐渐延长,可达5-10厘米并多少外弯或下弯,生有许多密集的小花;花序轴常较肥厚;花苞片卵状三角形或卵状披针形,长约2毫米,宿存,常稍呈紫色;花梗和子房长约3毫米;花黄绿色,平展,不甚张开;中萼片卵状披针形或长圆状披针形,长2-2.5毫米,下部宽约1毫米;侧萼片与中萼片相似,但背面具龙骨状突起或有时呈狭翅状;花瓣近长圆形或狭长圆形,长约2毫米,宽约0.7毫米;唇瓣卵状三角形,长约1.8毫米,基部宽约1.2毫米,先端渐尖,边缘内卷;蕊柱短,无蕊柱足;花药狭卵形,直立,长约0.7毫米;蕊喙较长,直立,长达1.2毫米。花期4-10月。
产台湾、海南和云南南部至东南部(景洪、勐腊、富宁)。生于溪谷旁树干上、山崖树枝上或林中石上,海拔1100米以下。尼泊尔、锡金、印度、缅甸、泰国、越南、马来西亚、印度尼西亚和菲律宾也有分布。模式标本采自菲律宾。
形态特征:假鳞茎聚生,扁球形(上下压扁),高3—10毫米,粗7—18毫米,顶端通常具1枚大叶和1(--2)枚小叶。大叶狭长圆状倒披针形至近狭长圆形,长(2.7--)4—8(一12)厘米,宽6—13毫米,先端钝、急尖或不等的2裂,稍肉质,基部收狭成短柄并内卷;小叶近长圆形,长0.7—1.5厘米,较少不存在。花葶生于假鳞茎基部,高10—20(--30)厘米,纤细,有2—3枚抱茎的鞘;总状花序初期较短,长1—2厘米,随着花的开放,逐渐延长,可达5--10厘米并多少外弯或下弯,生有许多密集的小花;花序轴常较肥厚;花苞片卵状三角形或卵状披针形,长约2毫米,宿存,常稍呈紫色;花梗和子房长约3毫米;花黄绿色,平展,不甚张开;中萼片卵状披针形或长圆状披针形,长2—2.5毫米,下部宽约1毫米;侧萼片与中萼片相似,但背面具龙骨状突起或有时呈狭翅状;花瓣近长圆形或狭长圆形,长约2毫米,宽约0.7毫米;唇瓣卵状三角形,长约1.8毫米,基部宽约1.2毫米,先端渐尖,边缘内卷;蕊柱短,无蕊柱足;花药狭卵形,直立,长约0.7毫米;蕊喙较长,直立,长达1.2毫米。花期4—10月。
产地分布:产台湾、海南和云南南部至东南部(景洪、勐腊、富宁)。生于溪谷旁树干上、山崖树枝上或林中石上,海拔1100米以下。尼泊尔、锡金、印度、缅甸、泰国、越南、马来西亚、印度尼西亚和菲律宾也有分布。模式标本采自菲律宾。
参考文献:Thelasis pygmaea(Griff.)Bl,F1.Jav.Orch.:23.1858;Lindl.In J.Linn.Soc.Bot.3:63.1859;Hance in J.Linn.Soc.Bot.13:127.1872;Hook.F., F1.Brit.Ind.6:86.1890;King et Pantl.In Ann.Bot.Gard.Calcutta8:249.P1. 331.1898;Seidenf.In Opera Bot.89:156.Fig.101.1986,et 114:193.1992;台 湾兰科植物彩色图鉴2:323,663,1990;2.H.TsietS.C.Chen in Acta Phytotax. Sin.33(3):296.1995;Z.H.Tsi et al,1,c.33(6):591.1995.——Eupro boscis Pygmaea Griff.In Calc.J.Nat.Hist.5:731.T.26,1845.——Thelasis triptera Rchb.F.In Bonplandia 3:219.1853;中国高等植物图鉴5:750. 图8329, 1976;海南植物志4:248.图1120.1977;台湾兰科植物3:272--273(图). 1987.——T.Elongata Bl,Bot.Mus.Lugd.2:187.1856.——T.honZAongen— sis Rolfe in Kew Bull.:199.1896,et in J.Linn.Soc.Bot.36:39.1903;S.Y, Hu,Gen.Orch.Hong Kong:fig.39.1977.——T.Clatisa Fukuyama inBot.Mag, Toky049:440.1935;Masamune in J.Geobot.21(1):t.187. 1973;台湾兰科植物 彩色图鉴1:504.1977;台湾植物志5:1109.1978.
本属概述:Bl,Bijdr.8:385.1825;Seidenf.In Opera Bot.89:154. 1986.——Euproboscis Griff. In Calc. J. Nat. Hist. 5:371. 1845.附生小草本,具假鳞茎或缩短的茎,后者常包藏于套叠的叶鞘中。叶1—2枚,生 于假鳞茎顶端或多枚二列地着生于缩短的茎上,后者基部具鞘并互相套迭,有时有关 节。花葶侧生于假鳞茎或短茎基部,通常较细长;总状花序或穗状花序具多花;花很 小,几乎不张开;萼片相似,靠合,仅先端分离;侧萼片背面常有龙骨状突起;花瓣略 小于萼片;唇瓣不裂,多少凹陷,着生于蕊柱基部;蕊柱短,无蕊柱足;花药直立,药 帽先端收狭为钻状;花粉团8个,每4个为一群,蜡质,共同连接于一个细长而上部稍 扩大的花粉团柄上;粘盘近狭椭圆形;蕊喙顶生,直立,渐尖,2裂(粘盘脱出后);柱 头较大。蒴果较小。本属模式种:Thelasis obtusa Bl.全属约20种,产亚洲热带地区,主要见于东南亚,向北可到尼泊尔和我国南部, 向南可到新几内亚岛。我国有2种。
亚族概述:Schltr,Orch.340.1914,etinNotizbl.Bot.Gard.Berlin9:576.1926."Thelasieae",附生草本,具或不具假鳞茎,有时有茎;茎一般纤细,长或短。叶茎生或生于假鳞茎顶端,扁平,对折,革质或近肉质,有时两侧压扁或近圆柱形,具或不具关节。花葶侧生;总状花序具少数或多数花;花一般较小,常不甚张开;萼片与花瓣离生,有时靠合;唇瓣生于蕊柱基部或短的蕊柱足上;蕊柱一般较短,具或不具蕊柱足;花药生于蕊柱顶端背侧;花粉团8或4个,蜡质,具有一个共同的花粉团柄,有粘盘;柱头1个,凹陷。本亚族模式属:Thelasis Bl.共含6属,分布于亚洲热带地区和大洋洲。我国有2属。
本族概述:Humb,Bonpl.EtKunth,Nov.Gen.Sp.Pl.1:269.1815.附生或较少地生或腐生草本,偶见攀援藤本;附生种类常有假鳞茎或肉质茎。叶基生或茎生,常为革质、纸质或肉质,罕有草质,少数无叶(腐生种类)。花变化较大;萼片合生或离生;唇瓣有距或无距;花药以狭窄基部或花丝连接于蕊柱,常俯倾,两药室一般密接并藏于药帽之内,花后常枯萎或脱落;花粉团一般4—8个,较少2个,蜡质或有时粒粉质,具或不具花粉团柄或粘盘,偶见粘盘柄;蕊喙一般位于柱头上方。本族模式属:EpidendrumL.共约340属,全球均有分布,主要见于热带。我国有29个亚族含72属。
亚科概述:地生、附生或腐生草本,具根状茎、块茎或假鳞茎。叶茎生或基生,折扇状、对折或具网状脉,草质至革质。花序(或花葶)顶生或侧生;花通常两侧对称,有特化的唇瓣,少有例外,一般扭转,具1室子房;蕊柱通常具1枚能育雄蕊;花粉粘合成团块,称花粉团;花粉团2--8个,粒粉质或蜡质,其一端常伸出花粉团柄并连接于粘盘,也有直接连接于粘盘或通过粘盘柄连接于粘盘的,较少既无花粉团柄又无粘盘柄与粘盘的;柱头1或2,凹陷或凸出;蕊喙通常存在,大或小。果实通常为蒴果;较少呈荚果状。种子通常两端有延长的翅,较少无翅并有厚的外种皮。本亚科模式属:OrchisL.共约690余属,全球均有分布。我国有4个族,42个亚族共167属。
本科概述:地生、附生或较少为腐生草本,极罕为攀援藤本;地生与腐生种类常有块茎或肥厚的根状茎,附生种类常有由茎的一部分膨大而成的肉质假鳞茎。叶基生或茎生,后者通常互生或生于假鳞茎顶端或近顶端处,扁平或有时圆柱形或两侧压扁,基部具或不具关节。花葶或花序顶生或侧生;花常排列成总状花序或圆锥花序,少有为缩短的头状花序或减退为单花,两性,通常两侧对称;花被片6,2轮;萼片离生或不同程度的合生;中央1枚花瓣的形态常有较大的特化,明显不同于2枚侧生花瓣,称唇瓣,唇瓣由于花(花梗和子房)作180°扭转或90°弯曲,常处于下方(远轴的一方);子房下位,1室,侧膜胎座,较少3室而具中轴胎座;除子房外整个雌雄蕊器官完全融合成柱状体,称蕊柱;蕊柱顶端一般具药床和1个花药,腹面有1个柱头穴,柱头与花药之间有1个舌状器官,称蕊喙(源自柱头上裂片),极罕具2—3枚花药(雄蕊)、2个隆起的柱头或不具蕊喙的;蕊柱基部有时向前下方延伸成足状,称蕊柱足,此时2枚侧萼片基部常着生于蕊柱足上,形成囊状结构,称萼囊;花粉通常粘合成团块,称花粉团,花粉团的一端常变成柄状物,称花粉团柄;花粉团柄连接于由蕊喙的一部分变成固态粘块即粘盘上,有时粘盘还有柄状附属物,称粘盘柄;花粉团、花粉团柄、粘盘柄和粘盘连接在一起,称花粉块,但有的花粉块不具花粉团柄或粘盘柄,有的不具粘盘而只有粘质团。果实通常为蒴果,较少呈荚果状,具极多种子。种子细小,无胚乳,种皮常在两端延长成翅状。科的模式属:OrchisL.全科约有700属20000种,产全球热带地区和亚热带地区,少数种类也见于温带地区。我国有171属1247种以及许多亚种、变种和变型。兰科是一个进化而复杂的科,至今对科内各类群间亲缘关系的了解仍然是十分初步的。在目前已发表的许多兰科系统中,以R.Dressier(1981,1993)系统较为全面,但该系统对鸟巢兰族(ridb.Neottieae)的处理是有欠缺的。幸而C.Seidenfaden等人在“TheOrehidsofIndochina”(OperaBot.114:5--502.1992)和“TheOrehidsofPeninsularMalaysiaandSingapore”(1992)中有较合理的处理。本书基本上采用了后者的框架,但将等级作了调整,而对亚族的组成则更多地参考Dressler系统(PhylogenyandClassifica-tionoftheOn:hidFamily,1993)。本书将兰科分为3个亚科,兰亚科之下分4个族。其中兰族(Orchideae)与万代兰族(Vandeae)是比较自然的类群,而鸟巢兰族(Neottieae)与树兰族(Epidendreae)则尚存在不少问题。在许多近代兰科系统中,上述的4个族均被作为亚科,在亚科之下又有大量的族,在族之下则是亚族。应当说,大多数亚族的建立是比较客观的。但族的组合则含有相当多的主观性,甚至是勉强的拼凑。因此,本书避繁就简,在兰亚科之下只承认4个族,族之下则包含较多的亚族。这样不仅简化了层次,而且更为客观和实用。当然,这也只是权宜之计,有些问题仍然未能解决,如鸟巢兰族与树兰族的特征交叉过多,有些亚族较为庞杂等。这里就不作详细讨论了。这里要说明的是,本书所提供的“亚科与族的检索表”和“兰亚科分亚族检索表”均系根据国产种类编写而成的,未必完全符合族与亚族的全面特征,在使用时务请加以注意。
亚科概述:地生、附生或腐生草本,具根状茎、块茎或假鳞茎。叶茎生或基生,折扇状、对折或具网状脉,草质至革质。花序(或花葶)顶生或侧生;花通常两侧对称,有特化的唇瓣,少有例外,一般扭转,具1室子房;蕊柱通常具1枚能育雄蕊;花粉粘合成团块,称花粉团;花粉团2--8个,粒粉质或蜡质,其一端常伸出花粉团柄并连接于粘盘,也有直接连接于粘盘或通过粘盘柄连接于粘盘的,较少既无花粉团柄又无粘盘柄与粘盘的;柱头1或2,凹陷或凸出;蕊喙通常存在,大或小。果实通常为蒴果;较少呈荚果状。种子通常两端有延长的翅,较少无翅并有厚的外种皮。本亚科模式属:OrchisL.共约690余属,全球均有分布。我国有4个族,42个亚族共167属。
本科概述:地生、附生或较少为腐生草本,极罕为攀援藤本;地生与腐生种类常有块茎或肥厚的根状茎,附生种类常有由茎的一部分膨大而成的肉质假鳞茎。叶基生或茎生,后者通常互生或生于假鳞茎顶端或近顶端处,扁平或有时圆柱形或两侧压扁,基部具或不具关节。花葶或花序顶生或侧生;花常排列成总状花序或圆锥花序,少有为缩短的头状花序或减退为单花,两性,通常两侧对称;花被片6,2轮;萼片离生或不同程度的合生;中央1枚花瓣的形态常有较大的特化,明显不同于2枚侧生花瓣,称唇瓣,唇瓣由于花(花梗和子房)作180°扭转或90°弯曲,常处于下方(远轴的一方);子房下位,1室,侧膜胎座,较少3室而具中轴胎座;除子房外整个雌雄蕊器官完全融合成柱状体,称蕊柱;蕊柱顶端一般具药床和1个花药,腹面有1个柱头穴,柱头与花药之间有1个舌状器官,称蕊喙(源自柱头上裂片),极罕具2—3枚花药(雄蕊)、2个隆起的柱头或不具蕊喙的;蕊柱基部有时向前下方延伸成足状,称蕊柱足,此时2枚侧萼片基部常着生于蕊柱足上,形成囊状结构,称萼囊;花粉通常粘合成团块,称花粉团,花粉团的一端常变成柄状物,称花粉团柄;花粉团柄连接于由蕊喙的一部分变成固态粘块即粘盘上,有时粘盘还有柄状附属物,称粘盘柄;花粉团、花粉团柄、粘盘柄和粘盘连接在一起,称花粉块,但有的花粉块不具花粉团柄或粘盘柄,有的不具粘盘而只有粘质团。果实通常为蒴果,较少呈荚果状,具极多种子。种子细小,无胚乳,种皮常在两端延长成翅状。科的模式属:OrchisL.全科约有700属20000种,产全球热带地区和亚热带地区,少数种类也见于温带地区。我国有171属1247种以及许多亚种、变种和变型。兰科是一个进化而复杂的科,至今对科内各类群间亲缘关系的了解仍然是十分初步的。在目前已发表的许多兰科系统中,以R.Dressier(1981,1993)系统较为全面,但该系统对鸟巢兰族(ridb.Neottieae)的处理是有欠缺的。幸而C.Seidenfaden等人在“TheOrehidsofIndochina”(OperaBot.114:5--502.1992)和“TheOrehidsofPeninsularMalaysiaandSingapore”(1992)中有较合理的处理。本书基本上采用了后者的框架,但将等级作了调整,而对亚族的组成则更多地参考Dressler系统(PhylogenyandClassifica-tionoftheOn:hidFamily,1993)。本书将兰科分为3个亚科,兰亚科之下分4个族。其中兰族(Orchideae)与万代兰族(Vandeae)是比较自然的类群,而鸟巢兰族(Neottieae)与树兰族(Epidendreae)则尚存在不少问题。在许多近代兰科系统中,上述的4个族均被作为亚科,在亚科之下又有大量的族,在族之下则是亚族。应当说,大多数亚族的建立是比较客观的。但族的组合则含有相当多的主观性,甚至是勉强的拼凑。因此,本书避繁就简,在兰亚科之下只承认4个族,族之下则包含较多的亚族。这样不仅简化了层次,而且更为客观和实用。当然,这也只是权宜之计,有些问题仍然未能解决,如鸟巢兰族与树兰族的特征交叉过多,有些亚族较为庞杂等。这里就不作详细讨论了。这里要说明的是,本书所提供的“亚科与族的检索表”和“兰亚科分亚族检索表”均系根据国产种类编写而成的,未必完全符合族与亚族的全面特征,在使用时务请加以注意。
《Flora of China》 Vol. 25 (2009)
Thelasis pygmaea
矮柱兰